
An Introduction to Tetrapods
The word Tetrapod comes from the Greek word Tetrapoda meaning four feet. Tetrapods are defined by having four legs and feet or descending from an animal that had four limbs. Today tetrapods encompass all land living-vertebrates such as amphibians, reptiles, birds, and mammals, as well as those animals such as seals and whales which returned to the sea and snakes which have lost limbs through evolution.
Evolution
Origins
The earliest tetrapod forms evolved at some stage during the Devonian. However, the specific ancestors and processes which led to the development of limbs and ultimately the colonisation of land remain uncertain. The adaptation to walk on the land is one of the most important evolutionary changes. It had profound impacts on life as we know it today and remains an active and exciting area of research in palaeontology.
Early tetrapods are believed to have evolved from lobe-finned fish, with Rhipidistians being the most likely ancestor due to the close anatomical similarities (Westoll, 1943). Rhipidistians were common in Devonian but became extinct during the Early Permian. The only lobe-fined fish that exist today are lungfish and coelacanths. Shared characteristics between Rhipidistians and early amphibians include a nearly identical skull, teeth, and vertebrate, the ability to breath air, and movable bones and muscles in ventral fins. The early adaptations for life on land evolved in the water before the transition to land had occurred. Lobe-finned fish may have initially evolved moveable fleshy ventral fins as a mechanism to improve swimming and to move about on the bottom of waters. The ability to breathe air had already developed in lobe-finned fish in the Devonian with initial lung-like structures originating as an outgrowth of the gut which may have developed to aid buoyancy as fish became increasingly armoured and heavy or to increase oxygen consumption.

Figure 1a. Reconstructions of Tiktaalik.
Tetrapodomorph fish, like Panderichthys and Tiktaalik, have been proposed as sister taxon to tetrapods (Ahlberg, 2018). Panderichthys and Tiktaalik retained lobe-finned fish characteristics such as paired fins, scales, gills, fish-like pelvis, the absence of sacrum, and had straight knees and elbows and posteriorly facing hips but began showing a tetrapod-like head and body shape. These forms were aquatic and walked along the bottom of shallow estuaries, perhaps occasionally pushing themselves out of the water to take advantage of low competition and predation risk food sources on land.

1b. Panderichthys rhombolepis
Over time, tetrapods adapted to become more suited to terrestrial environments. In order to complete the transition from the sea to land early tetrapods needed to make many anatomical adjustments. Moving from an aquatic to land based environment requires that organisms adapt to support their entire weight and mitigate dehydration. While tetrapodomorphs were on the path to obtaining some of these capabilities, tetrapods evolved further acquiring flexed elbows and knees, digits, a sacrum, laterally projecting shoulder and hip joints, and more developed vertebrates. Early tetrapod forms had crocodile-like heads, dentitions suitable for catching relatively large prey, raised eyes paired with an ability to keep heads above ground for better aerial vision and a movable neck, skeletal adaptations for better weight support, and more efficient substrate locomotion better suited for life in shallow water and on shorelines, and nostrils for breathing air (Ahlberg, 2018). However, all of these adaptations were not obtained at once making it difficult to distinguish between the most primitive tetrapods and the most advanced terapodomorphs.
The first well preserved tetrapod body fossils come from Greenland where whole body reconstructions for two species have been found. These are Ichthyostega and Acanthostega which both possess a mosaic of primitive and derived features (Jarvik, 1955; Clack 1988). These early tetrapods retained gills, had limbs that struggled to support their body weight, and hands and feet that were more fin than finger. Acanthostega, dating to 365 million years ago, was about 2 feet long, weighed 2 pounds, and had short limbs ending in 8 webbed digits and joints which were not weight-bearing, in addition to fish features such as lateral sensory line running down the body and the absence of a wrist structure. Acanthostega was likely completely aquatic with hind legs acting as paddles. Ichthyostega was about 5 feet long, weighed 50 pounds, had a narrow braincase broad overlapping ribs, large shoulders, and 7 digits. Ichthyostega was likely amphibian spending part of its time on land. While Acanthostega and Ichthyostega share some common features it is unlikely that they are closely related and both represent side branches of tetrapods which were ultimately evolutionarily unsuccessful.
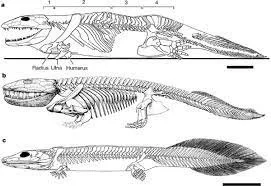
Figure 2. Reconstruction of Ichthyostega from Ahleberg et al. (2005).
In 2019 a tetrapod named Parmastega aelidae was discovered in a tropical lagoon environment in Russia (Beznosov et al., 2019). Over 100 well preserved bones have been uncovered from the head to shoulder girdle, of varying sizes, the maximum with a head length of 27cm and a body size of over a metre. These fossils have been dated to 372 million years ago, making them the oldest tetrapod recorded.This animal was likely a gill breather which retained fish-like characteristics in some bones and sensory organs for detecting vibrations in the water with a large skull, oval-shaped eyes which are positioned high towards the top of the skull, suggesting the animal looked above the surface
of the water but with a low external nasal opening which would have remained underwater. The retention of a shoulder girdle partly composed of cartilage suggests that this species never really left the water.
The Devonian body fossils record shows evidence of mosaic evolution with different species acquiring various adaptations for life on land at different times. Over 11 genera of tetrapod have been described from the Devonian, some of which are only known from jawbones and other fragmentary material. These genera would have varied on the degree to which they were aquatic or amphibian. The fossils record has a wide geographic distribution and diversity, with fossils found in Asia (Zhu et al., 2002), Russia (e.g. Lebedev & Coates, 2004; Lebedev, 2004), North America (Daeschler et al., 1994), Europe (Ahlberg, 1995, Ahlberg et al., 1994), and Australia (Young, 2006). All these fossils bar those from Australia which belong to the ancient continent of Gondwana were from the Laurussian supercontinent.
The first true terrestrial tetrapods were likely small with size increasing later as they mastered the land. Devonian tetrapods would have been primarily aquatic, remaining firmly tied to the water while expanding exploits on land. It is likely that fully terrestrial tetrapods did not arise until the Early Carboniferous.
Theories of Evolution
Much debate surrounds the environments from which tetrapods emerged and the environmental pressure which pushed them out of the water. Many environmental origins have been suggested including intertidal flats, brackish to freshwater vegetated areas (Niedzwiedzki et al., 2010; Blieck et al., 2007), flooded woodlands (Retallack, 2011), shallow-marine lagoons (Narkiewicz et al., 2015), and ephemeral lakes (Qvarnstorm et al., 2018). The earliest records appear from inland river basins, sabkha plains, and ephemeral coastal lakes, areas which tend to have poor fossil preservation. Some of the common environmental forces proposed are the drying up of lakes and rivers in an arid climate which favoured organisms that could withstand dry conditions and escaping predation in an ocean that was dominated by huge fish. The transition to shallow water and then to land made sense as not only was it a way to escape large predators on land but also gave an opportunity to prey on other smaller fish which had retreated to the shallows and eventually exploit new food sources on land. Other evolutionary drivers include estivation, larval competition, basking on land, and anoxia survival.
It is possible that tetrapod species evolved independently across these environments and in response to a number of different factors at various times, which makes it difficult to determine the single common ancestor of all modern tetrapods.
Early Tetrapod Evidence
Trace Fossils
The oldest widely accepted evidence of tetrapods comes from fossilised trackways on Valentia Island which were discovered by Ivan Stössel in 1992 (Stössel, 1995). These trackways formed around 385 million years ago when an early tetrapod dragged itself along an environment of muddy floodplains, lakes, and river channels around 200km from the sea (Stössel et al., 2016). The maximum size of this tetrapod was roughly 1m, similar to that of Ichthyostega. These trackways are contemporary with the earliest known Panderichthys, suggesting that at this point an evolutionary gap from forelimb to hindlimb propulsion had already been breached. The coexistence of tetrapods and tetrapodomorphs suggests that tetrapods may not have evolved directly from these forms. All recovered tetrapod body fossils are younger than the Valentia Island trackway.
Prior to the discovery of the Valentia trackway three Devonian trackways had been recorded, two in Victoria, Australia (Warrant et al., 1986; Warren & Wakefield, 1972), and one at Tarbat Ness, Scotland (Rogers, 1990). None of these trackways give a verifiable age older than the Valentia Island fossils.
In 2010, a tetrapod trackway with digit imprints dating to 397 million years ago was discovered at Zachelmie Quarry in Poland. This date makes them at least 10 million years older than the Valentia Island trackway and oldest than Panderichthys (Qvarnstrom et al., 2018). These tracks were found in what has been described at various stages as a tropical lagoon (Narkiewicz & Retallack, 2014; Narkiewicz et al., 2015), intertidal setting (Niedzwiedzki et al. 2010; Janvier and Clément, 2010), or shallow ephemeral lakes on a coastal plain (Qvarnstorm et al., 2018)). The animal that made these tracks has been estimated to have been 2.5m large, making it much larger than any other early tetrapod record (Nidezwiedzki et al., 2010; Clack, 2012). For many reasons, this discovery shook the palaeontology world by casting doubt on established evolutionary consensus. The discovery suggested that it was fish rather than amphibian that first developed limbs, questioning the longed held assumption of Panderichthys as a transitional form between fish and tetrapod and that the fish to tetrapod transition may have occurred as early as the Lower Devonian. The finding also pushes for a scenario in which tetrapod evolution occurred in a marine intertidal or lagoon environment rather than a freshwater

Figure 3. Tetrapod trackway at Dohilla, Valentia Island.
environment as suggested by the fossil record up to that point. However, the tetrapod origin of these prints has been contested and has been proposed to be fish feeding or resting traces instead (Lucas, 2015; 2019).
Share the Adventure
References
Ahlberg, P. E. (1995). Elginerpeton pancheni and the earliest tetrapod clade. Nature, 373, 420–425.
Ahlberg, P., Luksevics, E., & Lebedev, O. (1994). The first tetrapod finds from the Devonian (Upper Famennian) of Latvia. Philosophical Transactions of the Roval Society of London, B343, 303–328.
Ahlberg, P. E., Clack , J. A., & Blom , H. (2005). The axial skeleton of the Devonian tetrapod Ichthyostega. Nature, 137–140.
Ahlberg, P. (2018). Follow the footprints and mind the gaps: a new look at the origin of tetrapods. Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 1-23. doi:10.1017/S1755691018000695
Beznosov, P. A., Clack, J. A., Lukševičs, E., Ruta, M., & Ahlberg, P. (2019). Morphology of the earliest reconstructable tetrapod Parmastega aelidae. Nature, 574, 527–531.
Blieck, A., Clement, G., Blom, H., Lelievre, H., Luksevics, E., Streel, M., . . . Young, G. (2007). The biostratigraphical and palaeogeographical framework of the earliest diversification of tetrapods (Late Devonian). In R. Becker, & W. Kirchgasser (Eds.), Devonian events and correlation (Vol. 278, pp. 219–235). Geological Society of London Special Publication.
Clack, J. (1998). New material of the early tetrapod Acanthostega from the Upper Devonian of East Greenland. Palaeontology, 31, 699 724.
Clack, J. (2012). Gaining Ground: The Origin and Evolution of Tetrapods. Bloomfield: Indiana University Press.
Daeschler, E. B., Shubin, N. H., Thomson, K. S., & Amaral, W. W. (1994). A Devonian Tetrapod from North America. Science, 265(5172), 639-642.
Janvier, P., & Clement, G. (2010). Muddy tetrapod origins. Nature, 463, 40–41.
Jarvik, E. (1955). The Oldest Tetrapods and Their Forerunners. The Scientific Monthly, 80(3), 141-154.
Lebedev, O. A. (2004). A new tetrapod Jakubsonia livnensis from the Early Famennian (Devonian) of Russia and palaeoecological remarks on the Late Devonian tetrapod habitats. ACTA UNIVERSITATIS LATVIENSIS Acta Uniersitatis Latviensis, 679, 79-98.
Lebedev, O. A., & Coates, M. I. (1995). The postcranial skeleton of the Devonian tetrapod Tulerpeton curtum Lebedev. Zoological Journal of the Linnean Society, 114(3), 307-348. doi:10.1006/zjls.1995.0027
Lucas, S. G. (2015). Thinopus and a Critical Review of Devonian Tetrapod Footprints. Ichnos, 22(3), 136-154. doi:10.1080/10420940.2015.1063491
Lucas, S. G. (2019). An ichnological perspective on some major events of Paleozoic tetrapod evolution. Bollettino della Societa Paleontologica Italiana, 58(3), 223-266. doi:10.4435/BSPI.2019.20
Narkiewicz, M., Grabowski, J., Narkiewicz, K., Niedźwiedzki, G., Retallack, G. J., Szrek, P., & De Vleeschouwer, D. (2015). Palaeoenvironments of the Eifelian dolomites with earliest tetrapod trackways (Holy Cross Mountains, Poland). Palaeogeography, Palaeoclimatology, Palaeoecology, 420, 173-192. doi:10.1016/j.palaeo.2014.12.013
Narkiewicz, M., & Retallack, G. (2014). Dolomitic paleosols in the lagoonal tetrapod track-bearing succession of the Holy Cross Mountains (Middle Devonian, Poland). Sedimentary Geology, 299, 74–87.
Niedźwiedzki, G., Szrek, P., Narkiewicz, K., Narkiewicz, M., & Ahlberg, P. E. (2010). Tetrapod trackways from the early Middle Devonian period of Poland. Nature, 463, 43–48.
Qvarnström, M., Szrek, P., Ahlberg, P. E., & Niedźwiedzki, G. (2018). Non-marine palaeoenvironment associated to the earliest tetrapod tracksNon-marine palaeoenvironment associated to the earliest tetrapod tracks. Scientific Reports, 8:(1074 ), 1-10. doi:10.1038/s41598-018-19220-5
Retallack, G. J. (2011). Woodland Hypothesis for Devonian Tetrapod Evolution. The Journal of Geology, 119(3), 235-258. doi:10.1086/659144
Rogers, D. (1990). Probable tetrapod tracks rediscovered in the Devonian of N Scotland. Journal of the Geological Society, London, 147, 746–748.
Stössel, I. (1995). The discovery of a new Devonian tetrapod trackway in SW Ireland. Journal of the Geological Society, 152, 407-413. doi:10.1144/gsjgs.152.2.0407
Stössel, I., Williams, E. A., & Higgs, K. T. (2016). Ichnology and depositional environment of the Middle Devonian Valentia Island tetrapod trackways, south-west Ireland. Palaeogeography, Palaeoclimatology, Palaeoecology, 462, 16-40. doi:10.1016/j.palaeo.2016.08.033
Warren, J. W., & Wakefield, N. A. (1972). Trackways of Tetrapod Vertebrates from the Upper Devonian of Victoria, Australia. Nature, 238, 469–470.
Warren, A., Jupp, R., & Bolton, B. (1986). Earliest tetrapod trackway. Alcheringa, 183–186.
Westoll, T. S. (1943). The Origin of Tetrapods. Biological Reviews, 18(2), 78-98. doi:10.1111/j.1469-185X.1943.tb00289.x
Young, G. C. (2006). Biostratigraphic and biogeographic context for tetrapod origins during the Devonian: Australian evidence. Alcheringa: An Australasian Journal of Palaeontology, 30(1), 409-428. doi:10.1080/03115510609506875
Zhu, M., Ahlberg, P. E., Zhao, W., & Jia , L. (2002). First Devonian tetrapod from Asia. Nature, 420, 760–761.